 |
Behavior of real ants
Real ants are capable of finding shortest path from a food source to the nest
(Beckers, Deneubourg and Goss, 1992; Goss, Aron, Deneubourg and Pasteels, 1989)
without using visual cues (Hölldobler and Wilson, 1990).
Also, they are capable of adapting to changes in the environment, for example finding a new
shortest path once the old one is no longer feasible due to a new obstacle
(Beckers, Deneubourg and Goss, 1992; Goss, Aron, Deneubourg and Pasteels, 1989). Consider the following figure in which ants are moving on a straight line which connects a food source to the nest:

It is well-known that the main means used by ants to form and maintain the line is a pheromone trail. Ants deposit a certain amount of pheromone while walking, and each ant probabilistically prefers to follow a direction rich in pheromone rather than a poorer one. This elementary behavior of real ants can be used to explain how they can find the shortest path which reconnects a broken line after the sudden appearance of an unexpected obstacle has interrupted the initial path (see next figure).
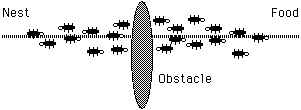
In fact, once the obstacle has appeared, those ants which are just in front of the obstacle cannot continue to follow the pheromone trail and therefore they have to choose between turning right or left. In this situation we can expect half the ants to choose to turn right and the other half to turn left. The very same situation can be found on the other side of the obstacle (see next figure).
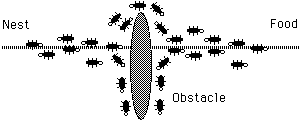
It is interesting to note that those ants which choose, by chance, the shorter path around the obstacle will more rapidly reconstitute the interrupted pheromone trail compared to those which choose the longer path. Hence, the shorter path will receive a higher amount of pheromone in the time unit and this will in turn cause a higher number of ants to choose the shorter path. Due to this positive feedback (autocatalytic) process, very soon all the ants will choose the shorter path (see next figure).
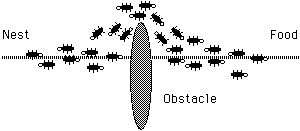
The most interesting aspect of this autocatalytic process is that finding the shortest path around the obstacle seems to be an emergent property of the interaction between the obstacle shape and ants distributed behavior: Although all ants move at approximately the same speed and deposit a pheromone trail at approximately the same rate, it is a fact that it takes longer to contour obstacles on their longer side than on their shorter side which makes the pheromone trail accumulate quicker on the shorter side. It is the ants preference for higher pheromone trail levels which makes this accumulation still quicker on the shorter path.
References
Beckers R., Deneubourg J.L. and S. Goss (1992). Trails and U-turns in the selection of the shortest path by the ant Lasius niger. Journal of theoretical biology, 159, 397-415.
Goss. S., Aron. S., Deneubourg J.L. and J.M. Pasteels (1989). Self-organized shortcuts in the Argentine ant. Naturwissenschaften 76, 579-581.
Hölldobler B. and E.O. Wilson (1990). The ants. Springer-Verlag, Berlin.
|
|



